
Conceitos e terminologias no estudo do ecossistema
- SPECO

- 6 de ago. de 2025
- 36 min de leitura
Atualizado: 11 de ago. de 2025
Por Maria Amélia Martins-Loução, Departamento de Biologia Vegetal, Faculdade de Ciências da Universidade de Lisboa, in Cadernos de Ecologia (3).
"Se o Todo-Poderoso me tivesse consultado antes de dar início à criação, teria recomendado algo mais simples." - Afonso X de Castela
Introdução
Ecologia, o estudo das relações dos organismos entre si e com o meio que os rodeia, é uma complexa e fascinante área da Biologia que tem enormes implicações para o Homem. A Ecologia não é intrinsecamente um campo de acção orientada; é antes uma área do conhecimento científico que trata o nível mais complexo de organização biológica. A ecologia pretende explicar porque é que os organismos se encontram num determinado ambiente e procura definir quais as variáveis físicas e biológicas que governam a sua distribuição.
Em ecologia podem considerar-se quatro níveis de organização: população, grupo de organismos da mesma espécie que se entrecruzam e ocupam uma dada área; comunidades, populações de diferentes organismos que vivem associadas no mesmo local; ecossistemas, comunidades em interacção com o meio que as suporta; biomas, conjuntos biológicos, caracterizados por determinadas formações vegetais, com comunidades animais uniformes, que ocorrem em grandes áreas geográficas, de características climáticas bem definidas. Dada a óbvia dependência entre espécies vegetais, animais e factores climáticos, cedo se reconheceu, nos alvores da ciência ecológica, ser o factor decisivo na diferenciação dos biomas e no estabelecimento e dinâmica das populações de organismos vivos, que lhe são próprios.
A ecologia, como ciência integradora, engloba um continuum de conceitos e escalas (Fig. 1) e grande diversidade de disciplinas tanto da biologia, geologia, hidrologia e meteorologia, como das ciências sociais e económicas.

O conceito ecologia foi introduzido pelo zoólogo alemão Ernst Haeckel, em 1866, com a seguinte definição: "por ecologia (do grego oekologia) entendemos a parte do saber respeitante à economia da natureza - a investigação das relações totais dos animais com o seu ambiente orgânico e inorgânico, incluindo acima de tudo as suas relações amigas e inimigas com os animais ou plantas com os quais venham a contactar directa ou indirectamente" (Deléage, 1991; Likens, 1992). Foi só, porém, no séc. XX que verdadeiramente se desenvolveu como ciência.
Se bem que o afloramento de temas e problemas ecológicos tenha sido contemporâneo da revolução industrial e da nova relação com o mundo que se instaura nas sociedades europeias a partir do final do século XVIII, é inegável que lhe encontramos já os traços, há mais de dois milénios, nos escritos de Aristóteles, Teofrasto e Plínio (Fig. 2).
Na sua mundivisão sobre a história natural, veiculada nas obras que chegaram até nós, pode perceber-se como estes autores se preocupavam e interrogavam sobre as relações entre os seres vivos e o ambiente, sem esquecer o papel específico do homem como factor no ordenamento e controlo da natureza.
Estabeleceram-se, desde a origem, dois pontos de vista paralelos e fecundos: o das populações de organismos, com a sua dinâmica específica, e o dos ciclos biogeoquímicos, que atravessam toda a biosfera e que vão dar origem a várias especialidades ecológicas ou subdivisões. Autonomizam-se, assim, a ecofisiologia, a ecologia das populações, a ecologia evolutiva, a ecologia aplicada, a ecologia vegetal, a ecologia animal, a fitossociologia, a paleoecologia. Estes diferentes campos de investigação ecológica podem ser agrupados em duas subdivisões: a autecologia, que estuda a reacção individual de cada organismo relativamente às variações do ambiente, e a sinecologia quando o objecto em estudo são populações ecomunidades.
Foi tendo em conta estas duas divisões que, actualmente, Likens (1992) propôs nova definição de ecologia: "estudo científico dos processos que influenciam a distribuição e abundância dos organismos, as interacções entre organismos e as interacções entre organismos e os fluxos de matéria e energia". Neste contexto, ecologia é o estudo das relações entre entidades ecólogicas, por exemplo organismos, populações, sistemas e o seu ambiente.
Até aos anos 60 o conhecimento e interesse pela ecologia era sobretudo científico. A partir do momento em que se dá mais importância aos problemas ambientais e às suas causas, alarga-se o interesse e geram-se expectativas em torno da ecologia, como ciência, capaz de resolver tais problemas ou pelo menos apontar possíveis soluções capazes de reabilitar os ecossistemas.
Em parte, a atenção crescente dada à ecologia, deve-se ao reconhecimento dos danos causados pelo homem sobre o ambiente, não só pelo crescimento urbano e industrial, de fortes consequências sócio-económicas (Fig. 3), mas também pelas fortes mudanças nas práticas agrícolas e florestais. Diversidade, abundância ou escassez de espécies existentes nos diversos ecossistemas, respectivos ciclos, sua estabilidade e regulação, tais são as questões que se põem à ecologia, em registo de crescente exigência e complexidade.

Devido à preocupação com os problemas ambientais que afectam a humanidade, a ecologia tem vindo a ser obrigada a responder a solicitações sociais precisas. Esta ciência torna-se, assim, inteiramente original pela forma como suscita debates de índole cultural, social e política, contra os efeitos do industrialismo no ambiente e na qualidade de vida. Como consequência, muitos cientistas, ao passarem a defender a responsabilização equitativa pelas carências de todos e a prudência ecológica, têm levado à confusão reinante entre a ecologia-ciência e a ecologia-política. Actualmente, assiste-se ao reforço da posição dos cientistas como profissionais da ecologia pelo papel activo que podem desenvolver na definição da política do ambiente,
A abordagem de qualquer problema ambiental é, no entanto, bastante complexa e quase sempre de difícil definição. Da mesma forma que a anatomia e a fisiologia se faz em níveis de integração que vão desde a célula à população, o objecto da ecologia começa no indivíduo e culmina na biosfera (Fig. 4).

A complexidade é ainda manifesta se se atende à escala espacial em que os processos e fenómenos ecológicos podem ser estudados, desde a escala molecular à escala do universo (Fig. 1 e 4). Embora a ecologia fosse - e ainda é - uma ciência empírica, a procura de conceitos unificadores aumento continuamente no séc. XIX, início do séc. XX, baseados em considerações teóricas, muitas vezes formalizados em termos matemáticos (Tamm, 1992). De entre esses termos, dois deles, inventados separadamente, ganharam larga aceitação: biogeocenose (Sukachaev, 1959) e ecossistema (Tansley, 1935). Embora considerados sinónimos, o termo ecossistema, usado muitas vezes também em sentido abstracto, foi considerado a unidade por excelência, por corresponder a um conceito holístico segundo o qual todos os seres vivos e o ambiente que os rodeia funcionam como um todo.
Conceito de Ecossistema
O conceito de ecossistema, criado pelo botânico inglês Tansley (1935), tem vindo a definir-se até hoje como um conceito fulcral no campo da ecologia. Anteriormente a Tansley a nomenclatura de unidades ecológicas, dotadas de maior ou menos autonomia, foi variada e ainda hoje é corrente o usod e termos como biocenose, microcosmos, holocenose ou biossistema. É apenas nos anos cinquenta, com Odum, que o conceito adquire a sua definição como unidade funcional (Odum, 1983).
A aceitação internacional deve-se certamente ao rápido crescimento da "análise de sistemas", como maneira de expressar as interacções entre componentes dum sistema, seja ele biológico, eléctrico ou mecânico (Aber & Melillo, 1991).
Um sistema pode ser definido como uma entidade integradora cujas propriedades gerais são diferentes das propriedades dos seus elementos. Neste sentido, um ecossistema constitui uma unidade em que, pela complexa interacção entre componentes bióticas e abióticas, se geram propriedades novas, quanto à estrutura e função, que ultrapassam a simples soma dos atributos pelas partes. Eles constituem sistemas abertos com graus de organização e auto-regulação, dependentes tanto do meio abiótico como dos organismos vivos, exercendo influência sobre o ambiente geral (Schulze & Zwölfer, 1987). De acordo com a nova escola de análise de sistemas (Margalef, 1968; Holling, 1973; Levin, 1978; Nicolis & Prigogine, 1989; Nicolis, 1991), os ecossistemas são, assim, considerados sistemas complexos com auto-organização: complexos por terem a capacidade de se adaptarem e ajustarem às variações das condições ambientais; com auto-organização porque as propriedades dos ecossistemas são o resultado das características das espécies que o constituem e não o resultado duma selecção directa (Solbrig, 1993; Schulze & Mooney, 1993).
Os ecossistemas são caracterizados pelos seus fluxos de energia, ciclos de carbono, água e iões e podem ser separados dos biomas pela escala e uniformidade do seu ambiente abiótico. Os ecossistemas são, pois, de interesse geral, porque a vida e evolução só são possíveis dentro do contexto destes sistemas. Este conceito aparece assim capaz de induzir uma convergência entre aspectos, fenómenos e procedimentos até agora desconexos, provenientes de diferentes campos científicos e incapazes, por si só, de originar modelos coerentes e globais do funcionamento da natureza que nos rodeia (Sarmiento, 1984).
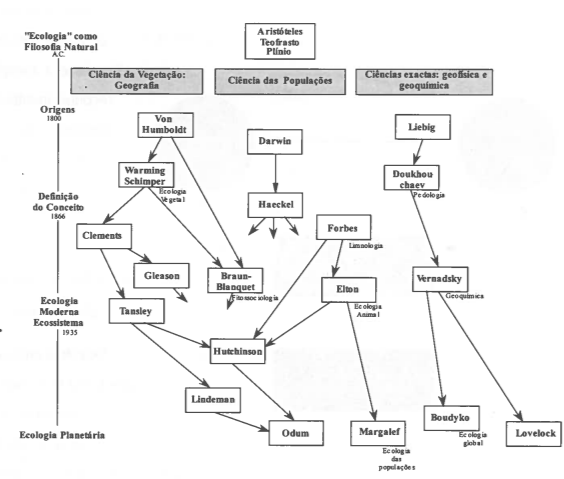
A dimensão dum ecossistema é muito variável. Tanto é um ecossistema uma floresta de coníferas, como um tronco de uma árvore apodrecido em que sobrevivem diversas populações de microflora e microfauna (Fig. 5). Assim como é possível associar todos os ecossistemas existentes num só, como um nível de organização muito elevado - a ecosfera, é igualmente possível delimitar em cada um, outros mais pequenos, por vezes ocupando áreas muito reduzidas que podem eventualmente receber o nome de microecossistemas (Fig. 6).

O ecossistema refere-se então a uma área, mais ou menos convencionalmente delimitada da ecosfera, considerada como um todo no espaço ou no tempo, onde se desenvolvem processos que vinculam os seus componentes distintos, bióticos ou abióticos, e que interactuam biunivocamente uns com os outros (Sarmiento, 1984). A forma de os limitar está em conhecer a importância relativa dos fluxos internos ou longitudinais de energia, relativamente aos fluxos laterais.
Os estudos ecológicos são normalmente realizados a diferentes escalas espaciais e temporais. Este facto dificulta a delimitação dum ecossistema como um todo porque, na maioria dos casos, a noção de limite ou fronteiras, não é, facilmente, considerada (Jarvis, 1987). Por esta razão, quando se usa o termo ecossistema no contexto de qualquer estudo ecológicos, os seus limites físicos e biológicos devem ser definidos com rigor (Moore & Chapman, 1986). No entanto, estes limites são, geralmente, determinados mais por razões práticas ou por conveniência do investigador do que pelo reconhecimento de descontinuidades funcionais com ecossistemas adjacentes (Likens, 1992). Em certa medida, pode dizer-se que tais limites dependem arbitrariamente dos objectivos do estudo.
Uma vez que a estrutura e função dos ecossistemas segue normalmente um padrão temporal, pode dizer-se que cada ecossistema tem uma história própria. Torna-se, por isso, importante, em especial para estudos comparativos, identificar o estádio de desenvolvimento do ecossistema e a sazonalidade da sua variação com o tempo (Likens, 1991).
No estudo dos ecossistemas são duas as principais abordagens:
1) análise dos fluxos de energia e matéria que neles têm lugar;
2) processos ecológicos e das mudanças que sofrem os indivíduos e populações que os integram.
Fluxos de Energia no Ecossistema
Definido como base na sua dinâmica, um ecossistema é um sistema aberto percorrido por fluxos contínuos de energia e matéria. Colectivamente, os organismos nos ecossistemas regulam a captação e gasto de energia e o ciclo dos minerais. Assim, a caracterização do fluxo de energia e dos ciclos biogeoquímicos e seus constrangimentos são essenciais para a compreensão do funcionamento e dinâmica das diferentes componentes de cada ecossistema.
A identificação precisa dos "inputs" e "outputs" dum sistema é o primeiro, senão o mais importante, passo para testar e analisar a sua complexidade. São estes contínuos inputs e outputs que estabelecem o contacto e permitem a transferência entre os diversos compartimentos do sistema, dando origem a ciclos que estão em equilíbrio no ecossistema, de acordo com as suas exigências e disponibilidade de recursos (Schulze & Mooney, 1993). A análise dos "inputs" e "outputs", em relação com o conhecimento das perturbações, pode revelar o grau de estabilidade ou variação dinâmica do sistema e ao mesmo tempo ajudar a esclarecer se as alterações são reversíveis ou irreversíveis. Por outro lado, a abordagem analítica pode ajudar a revelar a capacidade de acumulação dos diferentes reservatórios do ecossistema e a sua resiliência para absorver as variações impostas externamente, permitindo prever a sua estabilidade (Schulze & Zwölfer, 1987).
A necessidade de equilíbrio termodinâmico é provavelmente a premissa mais geral para definir as fronteiras dum ecossistema, uma vez que os diferentes fluxos, i.e. água, iões e energia, podem sofrer transformações químicas ou conversões, como no caso da energia.
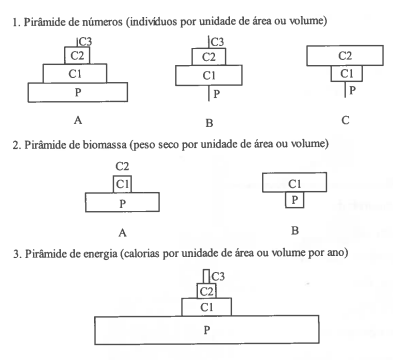
Relativamente à generalidade dos sistemas conhecidos, os ecossistemas possuem características particulares ligadas ao modo como neles se opera a utilização de energia. Do fluxo de energia que atinge cada ecossistema, apenas uma parte íntima é captada pelos organismos fotossintéticos - os organismos autotróficos ou produtores - capazes de transformar a energia radiante em energia química, utilizada posteriormente pelos organismos heterotróficos ou consumidores. Considerando um ecossistema como um todo, pode definir-se como produtividade primária bruta (GPP) a quantidade total de energia que é convertida em compostos orgânicos numa dada área e por unidade de tempo. A produtividade primária líquida (NPP) do ecossistema é a quantidade total de energia fixada por unidade de tempo menos a que é gasta durante as actividades metabólicas dos organismos na comunidade, i.e., a respiração (R).
GPP = NPP - R

Os organismos fotossintéticos, terrestres e aquáticos, fixam, em geral menos de 5% da energia solar que atinge os ecossistemas. Quando são consumidos, somente uma fracção da energia acumulada na biomassa, em média de 10%, pode ser convertida na biomassa dos organismos consumidores. De acordo com a 1ª lei da termodinâmica a energia nunca se perde apenas se transforma; ela entra no ecossistema sob a forma de energia solar e move-se dos produtores para os consumidores. Por causa dos gastos energéticos que em cada nível são requeridos para manutenção e crescimento, a energia que fica disponível para o nível seguinte é cada vez menor. Sendo a matéria orgânica produzida por unidade de tempo maior nos produtores, o rendimento da produção biológica diminui, assim, dos autotróficos para os heterotróficos. A Fig. 8 ilustra estas perdas de energia em cada nível numa cadeia trófica.

Dentro dos consumidores, podem ser considerados os consumidores primários - herbívoros - e os consumidores secundários - carnívoros. Outra componente importante da cadeia alimentar são os detritívoros ou microconsumidores que se alimentam de detritos ou de matéria orgânica vegetal ou animal. Há duas grandes classes nos detritívoros: os consumidores de detritos, essencialmente constituídos por pequenos animais e os decompositores, que consomem partículas de matéria orgânica em diferentes graus de decomposição e a transformam em inorgânica (Miller, 1990). Este grupo é essencialmente constituído por bactérias e fungos.
Todos estes tipos de organismos estão em maior ou menor grau representados no ecossistema, constituindo os níveis tróficos das cadeias alimentares. A complexidade de cada nível trófico varia muito segundo o tipo de ecossistema considerado.
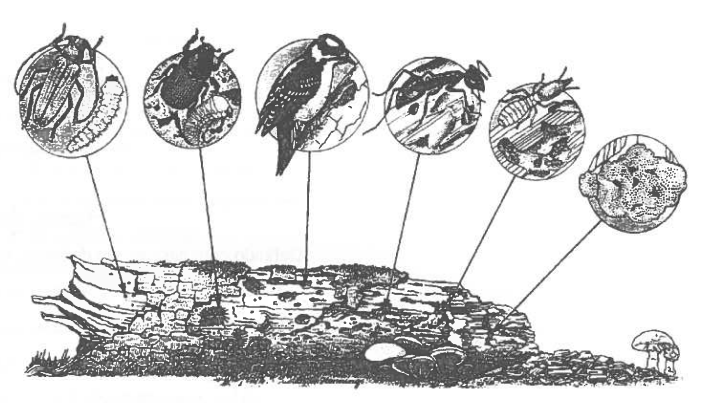
Estas relações, que se estabelecem devido à transferência de energia de nível trófico para nível trófico, podem ser graficamente representadas por meio de pirâmides ecológicas ou eltonianas. Deve-se a Charles Elton, em 1927, o conceito novo de pirâmide de números, com o intuito de simplificar o estudo das "sociedades animais" (Deléage, 1991).
No entanto, a profundidade do conceito eltoniano ganha definitivamente importância 20 anos mais tarde com o emergia da concepção moderna de ecossistema. Tais pirâmides podem ter expressão com base no número de indivíduos, na energia, ou menos na biomassa (Fig. 9). O primeiro nível representa a produção dos organismos autotróficos e os níveis superiores a produção dos herbívoros e carnívoros. A pirâmide de números para um ecossistema é decrescente, desde o nível dos produtores aos níveis tróficos mais elevados (Fig. 9.1.A). Mas, para outros ecossistemas, as pirâmides podem ter diferentes formas (Fig. 9.1.B), podendo ser, inclusivamente invertidas (Fig. 9.1.C). O peso total dos organismos vivos dum ecossistema é a biomassa, que aumenta à custa da produtividade primária líquida. Cada nível trófico, numa cadeia alimentar, contém uma certa quantidade de biomassa, que pode ser estimada colhendo amostras ao acaso ou seleccionando faixas representativas do ecossistema. Tal como o que se verifica para a maioria dos ecossistemas terrestres estas pirâmides são decrescentes (Fig. 9.1.A e 9.2.A), mas, nos ecossistemas aquáticos, podem ser invertidas (Fig. 9.1.B e 9.2.B).
Nestes casos, os produtores, que constituem o fitoplâncton, formado por microalgas, têm elevadas taxas de crescimento e reprodução, mas pequena biomassa. No entanto, a maneira simplificada de representar as pirâmides alimentares é bastante estática e não mostra as frequentes interferências externas nem o modo como o ecossistema é regulado e se ajusta a variações nos seus comportamentos bióticos e abióticos. Lindeman (1942), ao propôr a abordagem "trófico-dinâmica" em ecologia faz a primeira tentativa para medir as eficiências de transferências energéticas entre diferentes níveis tróficos animais. A eficiência ecológica, representada em diagrama na Fig. 10, refere-se ao rendimento segundo o qual os animais conseguem energia para sobreviver. A eficiência ecológica é por isso muitas vezes chamada eficiência de Lindeman.
Mais tarde, Dansereau (1980) passou a utilizar, ainda com base no conceito das pirâmides tróficas, a representação de uma esfera onde as interacções entre os diferentes níveis e o exterior eram tomadas em conta (Fig. 11).

Com esta esfera, Dansereau considerou outros níveis tróficos, partindo da base minerotrófica, que representa o solo nos ecossistemas terrestres ou a água nos aquáticos, atribuindo-lhe a importância que obviamente tem no ecossistema. À fitotrofia seguem-se dois níveis de zootrofia, correspondentes aos herbívoros e carnívoros e, pela primeira vez, dois outros níveis, o investimento e o controlo.
Estes dois últimos pretendem representar as quantidades de energia consumida quer no investimento dos diferentes níveis quer na regulação ou ajuste e adaptação do ecossistema a novas situações controlo incluindo a gestão nos ecossistemas humanos. Este tipo de representação contempla e realça ainda a corrente de energia que atravessa o ecossistema.
Existem ainda outras formas de representar diagramaticamente as interacções entre os diferentes níveis tróficos como por exemplo o de Moore & Chapman (1986), onde, igualmente, os "inputs" e "outputs" e as interacções com o meio exterior são tidos em conta (Fig. 12).
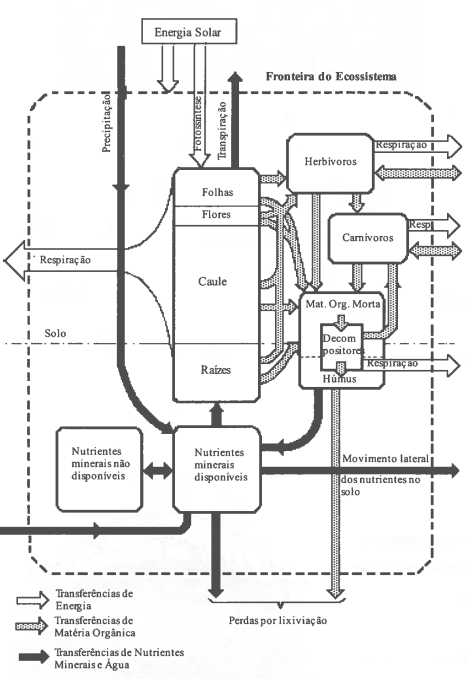
Muitas outras representações poderiam ser apresentadas sendo a mais simples aquela que considera que o ecossistema é apenas uma caixa negra com "inputs" e "outputs" de energia e matéria (Jarvis, 1987).
Ciclo de Nutrientes
O ciclo de nutrientes, inerente ao funcionamento dos ecossistemas e determinantes para a sua estrutura, é um mecanismo homeostático, no qual as plantas constituem um "pool" de nutrientes disponível ou trocável, incorporado sob a forma de biomassa viva ou morta. A acumulação deste "pool" através do tempo, acompanha a sucessão, enquanto que a sua redução acompanha a degradação do ecossistema.
Odum (1971) realça que o fluxo de nutrientes é mais significativo para o funcionamento dos ecossistemas que a sua concentração absoluta presente num dado compartimento e numa dada altura. A transferência entre compartimentos pode ocorrer através da tomada de nutrientes pelas plantas, lixiviação, decomposição biológica e troca entre o "pool" de nutrientes disponível, o solo e a própria rocha.
Todos os organismos vivos incorporam na biomassa elementos minerais que são parte integrante de diversos compostos orgânicos, com funções vitais no metabolismo, crescimento e reprodução. O ciclo de nutrientes no ecossistema é, pois, condição fundamental para a manutenção da vida, tanto dos autotróficos como dos heterotróficos. Alguns circulam entre o solo / água-organismos-atmosfera, outros circulam entre o solo-organismos-solo / água. Em qualquer dos casos, a abundância dum elemento particular na solução solo tem papel fundamental na distribuição das espécies, em particular dos vegetais. A distribuição dos organismos fica assim determinada pela interacção de certo número de factores, incluindo climáticos e biológicos, em conjunto com os edáficos. Estas interacções podem ser selectivas originando evoluções rápidas no ecossistema.
Na grande maioria dos ecossistemas a deficiência em certos nutrientes constitui, depois do factor hídrico, o factor ecológico limitante mais frequente. Os nutrientes que normalmente limitam a produtividade são o azoto e fósforo, podendo, em algumas circunstâncias, em ecossistemas continentais, incluir-se o potássio. Nestas condições, os organismos autotróficos desenvolveram estratégias evolutivas no sentido da conservação dos nutrientes através da reciclagem eficiente, com perdas mínimas. Nos ecossistemas podem, assim, existir diferentes tipos de processos, que se desenvolvem ao nível dos indivíduos, dos compartimentos bióticos ou ao nível do ecossistemas, como um todo (Fig. 13).

No primeiro caso (1) a reutilização dos nutrientes é contínua, com a tomada destes sem passagem pelos reservatórios do solo e folhada. No segundo (2) a recirculação faz-se sem passagem pelo solo mineral, ou seja os nutrientes são tomados directamente a partir da folhada em decomposição sem passar por uma fase inorgânica. Dados recentes de Chapin e colaboradores (1992) mostram que, em ecossistemas do tipo tundra, as plantas podem competir com os microorganismos pelo azoto orgânico. No terceiro nível, os ecossistemas conservam os nutrientes recirculando-os eficientemente entre o solo-planta-solo (3) ou os nutrientes são lixiviados pelas águas das chuvas (4), passando para a fase inorgânica do solo, através da mineralização microbiana e incorporados no "pool" de nutrientes acessível às plantas, ou ainda perdidos para a atmosfera, a partir da qual podem novamente entrar no ecossistema (Sarmiento, 1984).
O "input" de nutrientes no ecossistema pode ocorrer sob diversas formas e há problemas metodológicos específicos respeitante à avaliação das quantidades envolvidas. Tendo presente os principais elementos, a tabela 1 sumariza os processos mais importantes que contribuem para o "input" no ecossistema.
Tabela 1. Processos que contribuem para o "input" (x) de nutrientes nos ecossistemas (Rosen, 1990).
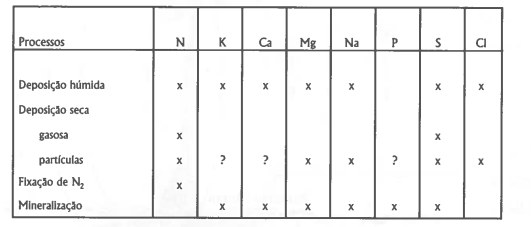
Geralmente, a sucessão duma comunidade pode ser deduzida do seu ciclo de nutrientes, através da análise dos "inputs" e "outputs". Estas medidas integram as reacções complexas de "turnover" dos nutrientes no ecossistema, daí o seu importante significado ecológico. Se os elementos estão a ser acumulados - "input" / "output" positivo, ou maior que 1 - nova biomassa está a ser acumulada e o sistema está a evoluir. Um "input" / "output" levemente negativo (ou menos que 1) pode representar mineralização da rocha mãe e sugere que o sistema está em equilíbrio (Barbour et al. 1980). A análise destes "inputs" - "outputs" dão informações acerca do "metabolismo" dos ecossistemas como se se tratassem de "super-organismos", podendo ser representados pela equação "Ulrich, 1987):
aCO2 + xM+ + yA- + (y-x)H+ + zH20 + Energia <--------->
(CaH2zOzMxAy)matéria orgânica + aO2
Fotossíntese e formação de matéria orgânica -------->
Respiração e mineralização de substâncias orgânicas <---------
em que a, x, y, z representam coeficientes estoiquiométricos, M+ catiões, A- aniões de valência unitária. Nesta equação, a "matéria orgânica" representa os produtores primários e secundários, i.e., o total de organismos, assim como o humus. Os catiões M+, aniões A-, e protões H+ estão presentes no solo, no "pool" de nutrientes acessíveis às plantas. Estas funções "metabólicas" operam entre as fronteiras dos sistemas, tais como a atmosfera, o solo mineral e a zona do solo circundante da raiz, a rizosfera.
Neste contexto, os iões presentes na solução solo e os iões de troca presentes na parte sólida do solo são aqui considerados como fazendo parte integrante do ecossistema (Bredmeier & Ulrich, 1992). Os catiões metálicos dos minerais, pelo contrário, são considerados componentes externos, uma vez que não estão imediatamente acessíveis para as reacções de transferência, pois têm que ser dissolvidos durante o processo de mineralização. Os iões lixiviados para zonas do solo abaixo das zonas das raízes, são considerados como "outputs" irreversíveis do ecossistema, o que é frequente, pelo menos em climas onde a precipitação excede a evapotranspiração. A análise dos "inputs"/"outputs", conjuntamente com a variação dos dados meteorológicos à escala espacial pode, assim, permitir a predição das tendências biogeoquímicas dos ecossistemas (Bredemeier & Ulrich, 1992).
Tanto a deposição atmosférica como os processos naturais do ciclo de nutrientes contribuem para a produção e consumo de H+, em especial nos ecossistemas florestais. O efeito destes fluxos no pH do solo depende das mudanças dos "pools" de H+ ao longo do tempo, e do equilíbrio dinâmico entre a capacidade tampão do solo e da quantidade de H+ livre na solução do solo. Embora o ciclo de nutrientes no ecossistema produza e consuma grandes quantidades de H+, estes ciclos equilibram-se não tendo, por isso, qualquer efeito na acidez do solo, a curto prazo. As tendências para a acidificação do solo a longo prazo são dominadas por transferências não cíclicas de H+ e de outros elementos, através das fronteiras do ecossistema, tais como a deposição atmosférica, produção de bicarbonatos, lixiviação e corte das florestas (Stevenson & Cole, 1999).
A recirculação de nutrientes parece ser o processo natural de fornecimento de nutrientes adequados às taxas de crescimento, assegurando deste modo a produtividade (Stevenson & Cole, 1999). No entanto, em muitos ecossistemas, há grandes limitações à tomada de nutrientes pelas plantas e à sua recirculação, como sejam infertilidade dos solos e baixas temperaturas, aliadas à presença de água no solo ou altas temperaturas em tempo seco. Nestas condições, as plantas terão de desenvolver estratégias adaptativas que lhes permita assegurar a sobrevivência e competição no ecossistema.
Intreracções entre Espécies
Os ecossistemas "funcionam" e evoluem devido à interacção complexa entre os seus organismos (Raven & Johnson, 1989). As interacções entre diferentes populações modificam o potencial genético de cada espécie (o seu óptimo fisiológico e a sua variação) de forma a permitir maior produção e manutenção a nível do ecossistema. As relações que se estabelecem regulam quer o fluxo de energia através dos ecossistemas, quer o ciclo de nutrientes e têm papel fulcral no funcionamento das comunidades.
As interacções estabelecidas no ecossistema, ou são positivas - como o comensalismo, em que um dos organismos beneficia e o outro não é afectado, ou o mutualismo, em que ambos beneficiam, ou negativas como o amensalismo, interacção que inibe o estabelecimento de outro organismo, ou a competição, interacção entre organismos que utilizam o mesmo recurso em condições limitantes.
Um caso típico de amensalismo é a interacção aleloquímica, em que há a inibição dum organismo por outro através da excreção de substâncias químicas para o meio ambiente. Estas substâncias tóxicas afectam somente algumas espécies. O significado ecológico destes aleloquímicos ainda não é verdadeiramente conhecido, embora desde sempre se tenha detectado que, dentro duma comunidade, se poderiam estabelecer ligações químicas entre organismos, decompositores, produtores e consumidores (Kormondy, 1996). As interacções aleloquímicas podem ser estabelecidas ao nível dos produtores-decompositores, ao nível dos produtores, ou ao nível dos produtores-consumidores.
Quando as interacções aleloquímicas envolvem apenas espécies vegetais o fenómeno chama-se alelopatia. Este fenómenos é especialmente importante e de elevado significado ecológico durante a sucessão da vegetação.
Ao longo do tempo, as características dos organismos coevoluem em relação uns aos outros, i.e. há ajustamentos dum grupo em relação a outro e vice-versa. A mais sofisticada e específica interacção química dá-se ao nível dos produtores-consumidores. Desde sempre as plantas desenvolveram adaptações morfofisiológicas contra a herbivoria. É o caso, por exemplo, do desenvolvimento de tricomas, tricomas glandulares, espinhos, esclerofilia, substâncias químicas, etc., que funcionam como defesas anti-herbivoria pela repulsa, pouca palatabilidade e fraca digestibilidade que criam. Deste modo, os herbívoros evitam comer certas plantas e selectivamente comme outras, ou adaptam-se e arranjam forma de rodear estas defesas. É interessante realçar que, quanto mais limitante é o habitat, mais especializações anti-herbívoria são desenvolvidas, e duma maneira geral, as plantas possuem mais do que um tipo de defesa.
As preferências dos herbívoros podem estar relacionadas não só com a palatabilidade como também por interacções hormonais. As interacções hormonais entre plantas e animais ocorrem a diferentes níveis e dependem da capacidade das substâncias químicas activas interagirem com diferentes tipos de organismos. O conhecimento das interacções hormonais entre plantas e animais veio de estudos entomológicos e da descoberta de que 1 ou 2 classes de hormonas de insectos estão presentes nas plantas (Harborne, 1992). A sua função é ainda especulativa, mas é possível que a sua deliberada produção tenha como objectivo a interferência na metamorfose dos insectos e, por isso, na sua reprodução. De entre as hormonas de insectos produzidos pelas plantas estão as ecdisonas que promovem o desenvolvimento das fases larvares. Estas hormonas, bastante frequentes nas plantas, com especial importância nos fetos e nas coníferas, assemelham-se às verdadeiras hormonas dos insectos produzidas pelo seu sistema endócrino e provocam uma contínua fase larvar impedindo, assim, que as pequenas larvas comam os vegetais ou originando, mesmo a inibição do desenvolvimento de gónadas maduras (Howe & Westley, 1988). Mais recentemente foi extraída do agerato (Ageratum houstonianum) uma hormona anti-juvenil, substância capaz de originar o desenvolvimento mais precoce das fases larvares dos insectos com a formação de insectos adultos estéreis, reduzindo, assim, o tempo de forte herbivoria por parte das larvas e ao mesmo tempo a reprodução dos insectos (Harborne, 1992).
As defesas anti-herbivoria mais importantes são as que envolvem produtos secundários do metabolismo, assim chamados para se distinguirem dos produtos primários que são regularmente formados como componentes dos principais processos metabólicos, tais como a respiração ou fotossíntese. Estes produtos, apesar de representarem gastos energéticos consideráveis, constituem investimentos importantes que asseguram a sobrevivência das plantas, em especial as que vivem em habitats pobres e de stress climático acentuado. Alguns destes produtos são sintetizados nas primeiras fases do desenvolvimento das plantas, são constitutivas; elas incluem a maioria das defesas mecânicas e alguns compostos químicos que reduzem a digestibilidade ou são mesmo tóxicos para os animais (tabela 2). Outros só são sintetizados quando necessário, como reacção imediata ao dano causado, são indutivas (Howe & Westley, 1988); por exemplo, as proteinases inibidoras ou outros produtos químicos, embora a lista possa vir a ser maior se se descobrir que muitas das toxinas até agora incluídas nas constitutivas, sejam efectivamente induzidas.
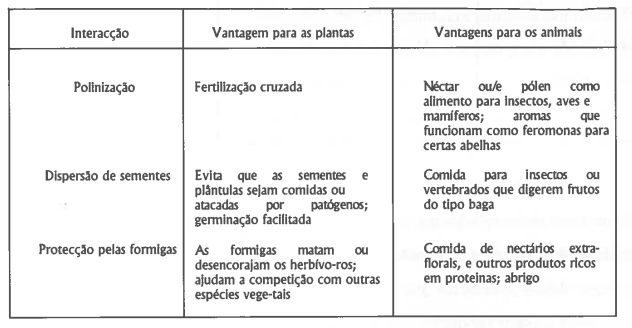
A herbivoria, consumo total ou parcial duma planta pelo consumidor é, assim, considerada por muitos autores como uma interacção negativa. No entanto, ela pode ter papel relevante na distribuição da vegetação e inclusivé estimular a produtividade. Por outro lado, é de realçar que existem interacções mutualistas não simbióticas de grande importância ecológica, entre herbívoros e plantas: a polinização, a dispersão de sementes e a protecção de plantas, realizada por determinadas espécies de formigas (tabela 3). Ao contrário da polinização e da dispersão das sementes, a defesa mutualista das formigas é bastante específica. Estas formigas, ao protegerem as plantas encontram não só néctar mas também abrigo e comida. Este tipo de adaptação anti-herbivoria por parte das plantas é o mais desenvolvido e interessante que se conhece, pois não envolve a produção de produtos secundários tóxicos (Howe & Westley, 1988).
Tabela 2. Produtos vegetais que reduzem a herbivoria. Adaptado de Howes & Westley (1988).
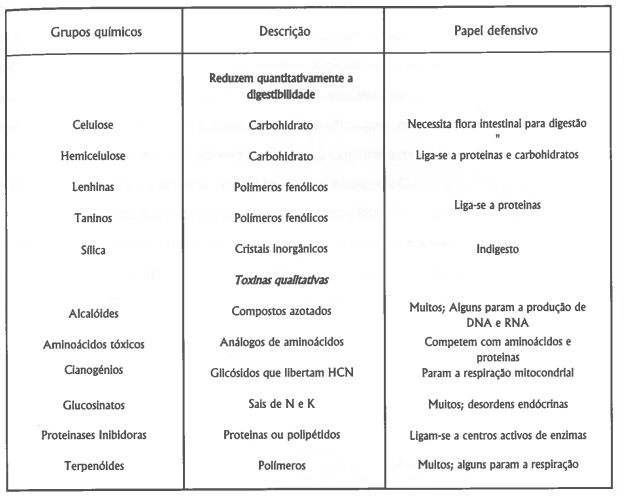
Tabela 3. Importantes mutualismos entre plantas e animais. Adaptado de Howes & Westley (1988).
Em resposta aos aleloquímicos, os herbívoros tornaram-se especializados e foram capazes de utilizar os produtos químicos obtidos nas plantas de que se alimentam para os utilizar contra os seus predadores. Noutros casos são capazes de os tornar e imobilizar para evitar os seus efeitos tóxicos. Muitos outros fenómenos tais como avisos, pigmentações de protecção ou mimetismo estão relacionados com a natureza dos químicos que os animais obtêm das plantas ou pelo seu próprio metabolismo. De facto, os animais, desde os insectos até aos vertebrados, também são capazes de fabricar os seus próprios produtos usando-os na defesa contra predadores. Estes animais, possuidores de substâncias tóxicas ou venenosas, desenvolvem pigmentações que servem de aviso aos predadores. Noutros casos, espécies desprotegidas são capazes, por mimetismo, de desenvolver cores semelhantes enganando, assim, os predadores. O mimetismo pode ser desenvolvido individualmente ou em espécies afins, originando um caso de defesa de grupo (Raven & Johnson, 1989).
As interacções positivas englobam-se nas relações simbióticas, definidas como relações em que dois ou mais organismos vivem sempre associados. Exemplos de simbioses são por exemplo os líquenes, as micorrizas, as associações de bactérias ou actinomicetes com raízes de certas plantas ou mesmo os bancos de corais, que envolvem a presença de animais e algas que, em conjunto, contribuem para a produtividade da associação. Alguns autores consideram que também o parasitismo é um caso de simbiose, uma vez que pressupõe a associação entre dois organismos. No entanto, neste caso, uma espécie beneficia da associação, enquanto a outra é prejudicada. Há também quem considere que o parasitismo é um caso de predação (Raven & Johnson, 1989). O comensalismo é um tipo de simbiose em que, na natureza, os indivíduos de uma espécie estão fisicamente ancorados aos indivíduos da outra. Por exemplo os epífitos, plantas que crescem nos ramos das outras, vários animais marinhos, como as lapas, que vivem em cima de grandes cetáceos e que assim são transportados de um lado para o outro, ou mesmo os pequenos peixes tropicais que vivem associados às anémonas (Raven & Johnson, 1989).
A exsudação de aleloquímicos ao nível das raízes dos vegetais, em especial na rizosfera, tem papel importante na selecção da população de microrganismos podendo actuar como selectores de determinadas associações simbióticas mutualistas ou como inibidores de parasitas ou mesmo patogénicos. A análise da ocorrência de associações mutualistas entre plantas e fungos ou bactérias nas comunidades naturais, mostra que são mais a norma do que a excepção. Mais de 90% das plantas formam associações com fungos formando micorrizas, com papel crucial para a tomada de fósforo e azoto. Grande número de espécies de Leguminosas têm bactérias fixadoras de N2 atmosférico do género Rhizobium como hospedeiros, formando nódulos radiculares. Por outro lado, muitas plantas lenhosas, arbustivas ou arbóreas, formam associações fixadoras de N2 com um actinomicete do género Frankia, dando origem a associações chamadas actinorrizas. Para além destas associações outras há em que as bactérias vivem em associação mutualista com raízes das plantas sem as penetrarem, mas intimamente ligadas, como é o caso de bactérias que fixam o N2 em vida livre, como o Azotobacter ou Azospirillum. Este tipo de associações é denominada rizocenose. As evidências fósseis das associações micorrízicas (Stubblefield et al., 1987) e fixadoras de N2 atmosférico (Sprent & Raven, 1985) sugerem que a selecção favoreceu fortemente este tipo de mutualismos (Read, 1993).
O impacto destes mutualismos na produção vegetal é proporcional à grandeza e natureza das limitações nutricionais a que as plantas estão sujeitas. Torna-se progressivamente maior à medida que a quantidade de azoto e fósforo decresce ao nível do solo. As mudanças das limitações nutricionais que se desenvolvem ao longo das sucessões primárias e secundárias leva a uma comparável sequência de comunidades dependentes de simbioses. Enquanto as plantas pioneiras da sucessão, que se estabelecem sem competição e na sua maioria em ecossistemas com baixo potencial de inóculo, são frequentemente não micorrizadas, elas são progressivamente substituídas por outras com hospedeiros (Read, 1993).
Nos ecossistemas naturais, os microorganismos responsáveis pelos ciclos biogeoquímicos do C, N, S e outros elementos, são "peças" importantes do ecossistema solo pois têm papel fundamental nos processos biogeoquímicos, formação do humus, transformação de compostos e energia. Como a sua identificação e caracterização envolve ainda problemas metodológicos são normalmente referidos não como organismos, mas antes como grupos fisiológicos. Daí que a ecologia microbiana não aborde a dinâmica de espécies microbianas mas sim de grupos funcionais, actualmente ainda muito pouco conhecidos, especialmente no que respeita à sua tolerância e sensibilidade aos factores ambientais e interacção com grupos funcionais de outros organismos (Meyer, 1993).
Também os parasitas têm capacidade para induzir marcadas mudanças na dinâmica das populações hospedeiras através da acção de forças selectivas que aumentam a mortalidade, reduzem a fecundação e diminuem a capacidade de competição. Experiências com plantas agrícolas têm mostrado os efeitos que nemátodos, fungos, bactérias e vírus têm nas plantas hospedeiras, mas estas informações não são suficientes para nos dar indicações acerca da sua importância no funcionamento dos ecossistemas naturais. Existem já evidências (Van der Putten, 2000) que mostram que os fungos parasitas são as forças selectivas que mais contribuem para a diversidade dos ecossistemas. Esta acção faz-se sentir a dois níveis: genético e estrutural. A nível genético os patogénicos afectam a diversidade intraespecífica e os padrões geográficos de distribuição da resistència, ao longo duma gama de espécies. Ao nível estrutural têm papel fundamental na fragmentação das populações e na diversidade das comunidades (Van der Putten, 2000). Neste contexto, não se pode excluir a sua acção no funcionamento e dinâmica dos ecossistemas, especialmente nas fases pioneiras e intermediárias da sucessão.
As interacções de espécies acabam por constituir um continuum com muitos casos intermediários, por exemplo entre o comensalismo e o parasitismo, mas de tal forma importantes que são capazes de determinar o padrão de distribuição das espécies nos ecossistemas. O conhecimento de todas estas relações é a base da compreensão dos fenómenos complexos que caracterizam a dinâmica do ecossistema.
Dinâmica e Funcionamento nos Ecossistemas
Os ecossistemas são sistemas abertos que funcionam como uma unidade ecológica, definida fundamentalmente pela sua dinâmica. À medida que o ecossistema amadurece passa a haver um aumento da sua biomassa total mas uma diminuição da sua produtividade líquida. Este processo gradual faz parte da tendência geral que os ecossistemas possuem para evoluírem ao longo do tempo, desde sistemas simples a outros mais complexos.
Durante muito tempo, em ecologia considerou-se que os ecossistemas mais complexos eram também os mais estáveis. No entanto há um continuum de forças que podem actuar num sistema, capazes de originar pequenas limitações à formação de biomassa - stress - ou de destruir total ou parcialmente a biomassa - perturbação. A inércia, resistência que um sistema oferece à mudança em condições de perturbação, é diferente da resiliência que diz respeito à capacidade dum sistema recuperar as suas características originais após perturbação (Dell et al., 1986).
A teoria pressupõe que os sistemas que possuem diferente número de espécies e diferentes interacções poderão diferir em estabilidade, variabilidade e resiliência. As populações podem apresentar menor variabilidade (flutuam menos) nas cadeias alimentares ricas em espécies com muitas ligações tróficas alternativas, porque exploram grande abundância de recursos. Isto equivale a dizer que, nos sistemas com grande biodiversidade, há pouca redundância porque o elevado número de espécies serve para diluir as variações que surjam. Pelo contrário, sistemas com baixa diversidade têm menor capacidade tampão quando há desaparecimento de espécies (Lawton & Brown, 1993).
De acordo com o modelo de Erlich & Erlich (1981), todas as espécies têm papel relevante na integridade da Biosfera, de modo pequeno mas significante. Daí haver limites relativamente ao número de espécies que podem desaparecer ou ser elimiandas antes do sistema colapsar. Nesta hipótese, chamada "hipótese do rebite" ou "rivet hypothesis", os autores comparam as espécies a rebites na estrutura dum avião. Um ecossistema é constituído por várias espécies que interagem e funcionam dando estabilidade ao sistema. Mesmo sob diferentes pressões elas podem funcionar como um todo e apresentar resiliência. De igual forma, num avião, os rebites da estrutura, apesar de sujeitos a diferentes forças estão colocados de modo a não saírem; a perda de um, por exemplo na asa do avião, pode eclodir num acidente grave. De acordo com esta teoria, um sistema biológico só responderia de forma catastrófica; o que não é o caso, uma vez que no sistema existem mecanismos de adaptação. As espécies não são "colocadas" no sistema como os rebites no avião; elas competem e estabelecem-se nos habitats adequados ao seu crescimento. Depois de instaladas podem modificar as condições do ecossistema, proporcionando o estabelecimento de outras espécies. Nesta sucessão de espécies e comunidades surgem "feedbacks" provocados pela limitação de recursos, que têm efeito estabilizador no ecossistema (Hobble et al., 1993).
A redução da biodiversidade é hoje acelerada pela destruição de habitats naturais. Esta perda merece preocupação porque a biodiversidade é uma "biblioteca genética" mantida pelos ecossistemas naturais, que nos fornecem "serviços" ainda muito pouco valorizados, tais como: controlo do clima regional, formação e manutenção do solo e ciclos biogeoquímicos, manutenção da composição atmosférica e controlo de pragas (Erlich & Wilson, 1991; Constanza et al., 1997). Neste contexto as espécies e a diversidade genética são críticas e fundamentais para se poder continuar a usufruir do benefício que os ecossistemas nos dão (Constanza et al., 1997).
As comunidades não são conjuntos de espécies ao acaso, nem mesmo nos sistemas aquáticos (Erlich, 1993; Steinberg & Geller, 1993). São antes constituídas por espécies com estreitas ligações funcionais através das quais se podem estabelecer relações mais gerais entre forma e função, desde que suficientemente estudadas e compreendidas a diferentes níveis do ecossistema (Körner, 1993). As correlações estabelecidas, com base em características de espécies vegetais funcionalmente agrupadas, oferecem a possibilidade de predição de processos ecológicos importantes. É neste contexto que surge a noção de espécies-chave ou essenciais (Bond, 1993) que podem ser olhadas como grupos funcionais sem representação redundante. Elas são chaves ou essenciais porque exercem um controlo desproporcional dentro do ecossistema, podendo originar mudanças acentuadas na estrutura das comunidades e no funcionamento dos ecossistemas, se eliminadas (Bond, 1993). Este tipo de espécies, de grande impacto no ecossistema, aparecem em todos os grupos desde os produtores aos consumidores e decompositores. Espécies que limitem a regeneração duma espécie dominante na comunidade ou espécies que afectam o regime de perturbação ou estabilidade de recursos podem ser consideradas neste grupo (Schulze & Mooney, 1993). Elas são importantes para a manutenção da integridade e estabilidade dos ecossistemas.
Os cientistas cedo reconheceram que a diversidade é importante para a variabilidade e manutenção dos processos ecológicos e para a resistência a mudanças externas. A noção de perda de diversidade é muitas vezes acompanhada por teorias que defendem que esta perda ameaça a integridade ecológica, o ambiente se torna frágil ou que a estabilidade do ecossistema fica afectada. A perda de espécies arrasta menor complexidade do sistema e consequentemente perda de estabilidade. Há, no entanto, larga controvérsia entre estabilidade e complexidade devido aos diferentes significados que estas palavras podem ter (Pimm, 1984). Muitos estudos analisam a estabilidade a diferentes niveis da organização ecológica, daí discutir-se a estabilidade de populações, composição de comunidades, ou biomassa das comunidades. O termo estabilidade é também muitas vezes relacionado com as diferentes estruturas das comunidades, ou sua complexidade, no seu sentido mais reducionista (Westman, 1986). Complexidade está associada ao número de espécies, ao grau de ligações interespecíficas ou à abundância relativa das espécies dentro das comunidades (Pimm, 1993).
Actualmente, a estabilidade pode ser caracterizada pela capacidade de resiliência e variabilidade das populações (Pimm, 1993). Uma alta resiliência é sinónimo de estabilidade, i.e., quanto mais estável é um sistema mais rápido retoma as suas características após uma perturbação. No entanto, a capacidade de resiliência depende de vários factores, entre eles a complexidade do sistema, ou seja a densidade de espécies e suas interacções. Por exemplo, quando há perturbação num grupo de espécies a sua capacidade de resiliência depende da resiliência das outras espécies com que estão interligadas. A variabilidade dum sistema depende também de um certo número de factores, entre eles a variabilidade espacial das populações e os fenómenos estocásticos (Westman, 1986; Pimm, 1993). É maior nos sistemas sujeitos a variações críticas das variáveis ambientais e depende da resiliência e da estrutura dos ecossistemas. Por exemplo, o aumento da diversidade de predadores pode reduzir a variabilidade de espécies sobre as quais eles exercem a sua acção. Neste contexto, a menor complexidade dos sistemas pode levar a uma maior variabilidade e resiliência (Pimm, 1993).
Os ecossistemas mediterrânicos, cuja evolução e dinamismo são o resultado duma acção directa do Homem, são dos ecossistemas que apresentam maior resiliência. Por essa razão, o estudo deste processo permite a previsão das respostas de tais sistemas à variedade de perturbações impostas pelo Homem. A resiliência constitui, assim, uma ponte entre a teoria e a aplicação em ecologia, permitindo focar os aspectos indutivos da ciência (Grubb & Hopkins, 1986).
O impacte ambiental da exploração do ecossistema tem implicações económicas e éticas, nomeadamente no que respeita à perda da diversidade genética e à mudança climática. Embora tendo presente a noção de produto cada vez se assiste mais à necessidade do equilíbrio sustentável do sistema, que é relevante para o funcionamento do ecossistema global. O objectivo mais importante é estabelecer o compromisso entre a necessidade de manter a produção ao longo do tempo e a protecção e conservação do ambiente, mantendo o uso eficiente dos recursos disponíveis. Para o ecólogo, o conceito de ecossistema sustentável, embora ainda pouco compreendido, pode trazer alguma informação acerca da resposta dos ecossistemas à mudança climática global.
A Biosfera e a Mudança Climática Global
O reconhecimento de que o homem tem papel fulcral na mudança climática global tem sido um dos aspectos mais significativos das últimas décadas. De facto, tomou-se consciência de que a actividade humana e muito especialmente os processos que contribuem para o aumento de gases na atmosfera - têm a potencialidade de trazer alterações a longo termo, por vezes irreversíveis, no nosso ambiente físico e biológico. Os efeitos destas mudanças físicas e químicas na atmosfera, nos fluxos de energia, no clima global e no nível das águas do mar estão interligados por fenómenos biológicos à escala global duma forma ainda mal compreendida.
Uma alteração no uso da terra é reconhecido no Programa Internacional da Geosfera e Biosfera (IGBP) (http://www.igbp.kva.se), como um dos factos mais importantes na mudança climática global, mas requer ainda grande investigação. A tradicional agricultura de abandono ganhou má reputação, porque a limpeza de novas áreas de terreno para a agricultura se faz através das práticas de corte e queima, provocando fortes emissões de CO2 para a atmosfera. Igualmente, a intensificação agrícola leva a grandes "outputs" de CO2 provenientes da oxidação da matéria orgânica. Sistemas sustentáveis parecem ser aqueles que minimizam os efeitos deletérios sobre o ambiente, ao mesmo tempo que asseguram as produções requeridas pelos agricultores (Swift & Anderson, 1993).
Recentes estudos têm mostrado que a vulnerabilidade dos sistemas agrícolas à mudança climática global parece apresentar disparidade entre países desenvolvidos e subdesenvolvidos. Testando as condições climáticas globais em diferentes zonas do globo, incluindo os efeitos combinados da mudança climática global e o aumento do CO2 na produção e na eficiência do uso de água, verificou-se que a produção agrícola diminuiria moderadamente nas latitudes mais baixas, onde se encontram os países mais subdesenvolvidos, e aumentaria consideravelmente nos países mais desenvolvidos (Rosenzweig & Parry, 1994). Por outro lado, simulando os cenários da mudança climática global de acordo com os dados do IPCC (Intergovernmental Panel on Climate Change) (http://www.ipcc.ch) verificam-se efeitos moderadamente negativos na produção cerealífera mundial, mesmo quando os efeitos directos do CO2, nível de desenvolvimento das explorações agrícolas e desenvolvimento tecnológico futuro são tomados em consideração. A única possibilidade de aumentar a produção é a instalação de rega o que, obviamente, acarreta custos económicos e ambientais difíceis de contabilizar (Rosenzweig & Parry, 1994). Os resultados destes estudos realçam, assim, implicações consideráveis para a futura distribuição mundial dos recursos alimentares e para o aumento da vulnerabilidade de países subdesenvolvidos. Mostram ainda a necessidade de estudos interdisciplinares no sentido de aumentar os conhecimentos acerca das interacções biológicas e sócio-económicas que podem resultar da mudança climática global.
Muito menos se sabe, porém, dos efeitos da interacção do aumento de CO2, temperatura elevada e stress hídrico sobre as comunidades naturais. Inicialmente, vão provavelmente afectar a variabilidade genética ao nível das populações, provocando a sua fragmentação e diminuindo a diversidade das espécies e paisagens. As espécies presentes na vegetação natural podem não estar adaptadas a estas novas combinações de constrangimentos ambientais derivados da mudança climática global ou podem encontrar-se tão longe dos locais mais adequados à sua sobrevivência que ficam impossibilitadas de os colonizar (McNaughton, 1993). As comunidades daí resultantes deverão ser pobres em espécies e altamente susceptíveis à invasão de infestantes, herbívoros, patogénicos e predadores que, em termos ecológicos, poderão restaurar a diversidade, até certo ponto (Schulze & Mooney, 1993). O gradual depauperamento da flora e da fauna pode, eventualmente, levar a certos tipos de ruptura de funcionamento dos ecossistemas, com os que foram encontrados em investigações teóricas de modelação ecológica (McNaughton, 1993; Hobbie et al., 1993). Há evidências de que a diversidade biótica quer em termos de diversidade genética de populações, quer em termos de diversidade de habitats e paisagens é crítica para a manutenção dos ecossistemas naturais e agrícolas (Myers et al., 2000; Pimm & Raven, 2000).
Existe ainda outro processo biológico que ameaça igualmente a mudança global da biosfera, provavelmente com mais impacte do que qualquer outro, o crescimento da população mundial. De facto, não há dúvidas de que o crescimento da população mundial é mais catastrófico do que qualquer antecipação ou predição da mudança climática global a curto prazo. Para além dos resultados directos da expansão urbana, o crescimento da população mundial é por si a força motriz que leva ao corte das florestas para aumentar a área de produção, e ao aumento das necessidades de energias não renováveis. A solução científica para o crescimento da população está já estabelecida, mas factores sociológicos, políticos e económicos têm papel crucial na resolução ou no adiar deste problema (Chaloner, 1991). O efeito do crescimento da população sobre a sociedade humana foi já sentido mesmo antes que os aspectos da mudança climática global se tornassem claros.
Conclusões
Neste texto procurou-se evidenciar os diferentes aspectos que controlam e regulam o funcionamento dos ecossistemas, dando realce, na parte final, ao potencial impacte da mudança climática global no fornecimento de alimento a nível mundial e na transformação das paisagens, em geral. Verifica-se, assim, a necessidade duma posição holística de conservação e recuperação dos ecossistemas de forma a manter-se a diversidade biológica e a produtividade dos ecossistemas como um todo. Esta posição deve ser baseada numa coordenada e eficiente educação ambiental. O verdadeiro nível de atenção da ecologia é o ecossistema global. Muitas vezes investe-se em esforços inúteis de análise de aspectos excessivamente parciais que estão sob o controlo de outros mecanismos que se ignoram completamente porque só se reconhecem e operam a níveis mais amplos, que não se presta atenção.
Para se alcançar um estudo completo do ecossistema há que desenvolver investigação interdisciplinar. No entanto, na maioria dos casos, a abordagem é feita mais sob a forma multidisciplinar sem interacção entre si. Isto deve-se ao facto dos termos multidisciplinar e interdisciplinar serem usados indiscriminadamente sem perfeita distinção. O objectivo é a interdisciplinariedade mas, a realidade, é a multidisciplinariedade.
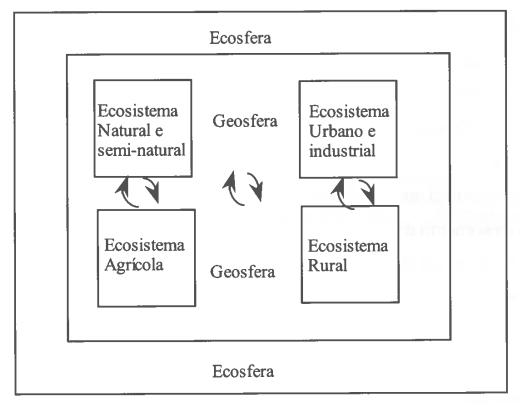
O conceito de ecossistema tem grande valor e importância per se, quer do ponto de vista político no ordenamento do território, quer do ponto de vista científico no desenvolvimento da compreensão das mudanças globais impostas pelo homem. Torna-se, por isso, cada vez mais importante desenvolver um verdadeiro esforço interdisciplinar no estudo do comportamento e dinâmica dos ecossistemas para se poder criar um planeamento e ordenamento integrado da ecosfera (Fig. 14), mais elevado nível de organização, que engloba a biosfera, a tecnosfera e a geosfera (Naveh, 1987).
Referências
Aber, J.D. & Melillo, J.M, 1991. Terrestrial Ecosystems. Sounders Publishing. Philadelphia, Tokyo. ISBN 0-03-047443-4
Barbour, M.G., Burk, J.H. & Pitts, W.D. 1980. Terrestrial Plant Ecology. The Benjamin / Cummings Publishing Company, Inc.
Bredemeier, M., Ulrich, B. 1992. Input / output - Analysis of Ions in Forest Ecosystems. In: Responses of Forest Ecosystems to Environmental Changes. Teller, A., Mathy, P. & Jeffers, J.N.R. (eds.). Elsevier Applied Science. pp: 229-243.
Chaloner, W.G. 1991. Global Change and The Biosphere: Introduction. Annals of Bot. 67 (Suplement 1): 1-3.
Chapin, F.S. III, Moilanen, L. & Kielland, K. 1992. Preferential use of organic nitrogen for growth by a non-mycorrhizal artic sedge. Nature 361: 150-153.
Constanza, R., D'Arge R., de Groot, R., Farber, S., Grasso, M., Hannon, B., Limburg, K., Naeem, S., O'Neill, R.V., Paruelo, J., Raskin, R.G. Sutton, P & van der Belt, M. 1997. The value of world's ecosystem services and natural capital. Nature 387: 253-260.
Dansereau, P. 1980. Ecologia Aplicada ao Ordenamento. Centro de Engenharia Biológica. Centro de Estudos Geográficos. Catarino, F.M., Daveau, S., Pereira, A.R., Cabral de Mello, A., Correia, A.I. (eds.)
Deléage, J.P. 1991. História da Ecologia. Uma Ciência do Homem e da Natureza. Puglicações D. Quixote. Nova Enciclopédia.
Dell, B., Hopkins, A.J.M., Lamont, B.B. 1986. Resilience in mediterranean-type ecosystems. Junk Publishers. Dordrecht.
Ehrlich, P.R. & Erlich, A.H. 1981. Extinction. The causes and the consequences of the disappearance of species. Random House, N.Y.
Ehrlich, P.R. & Wilson, E.O. 1991. Biodiversity Studies: Science and Policy. Science 253: 758-762.
Ehrlich, P.R. 1993. Foreword. Biodiversity and Ecosystem Function: Need We Know More? In: Biodiversity and Ecosystem Function. Schulze E.D. & Mooney H.A. (eds.) Ecological Studies 99. Springer Verlag. Berlin. pp: VI-XI.
Grubb, P.J. & Hopkins, A.J.M. 1986. Resilience at the level of the plant community. In: Resilience in mediterranean-type ecosystems. Dell, B. Hopkins, A.J.M., Lamont, B.B. (eds.). Junk Publishers. Dordrecht.
Harborne, J.B. 1992. Introduction to Ecological Biochemistry. 4th Edition. Academic Press. London.
Hobbie, S.E., Jensen, D.B. & Chapin III, F.S. 1993. Resource Supply and Disturbance as Controls over Present and Future Plant Diversity. In: Biodiversity and Ecosystem Function. Schulze E.D. & Mooney H.A. (eds.) Ecological Studies 99. Springer Verlag. Berlin. pp: 385-408.
Holling, S.C. 1973. Resilience and stability of ecological systems. Annu. Rev. Ecol. Syst. 4:1-24.
Howe, H.F. & Westley, L.C. 1988. Ecological Relationships of Plants and Animals. Oxford University Press. N.Y., Oxford.
Jarvis, P.G. 1987. Water and carbon fluxes in ecosystems. In: Potentials and Limitations of Ecosystem Analysis. Schulze E.D. & Zolfer H. (ed.) Ecological Studies 61. Springer Verlag. Berlin.
Kormondy, E.J. 1996. Concepts of Ecology, 4th Edition. Prenctice Hall Publishers. N.J.
Korner, Ch. 1993. Scalling from Species to Vegetation: The Usefulness of Functional Groups. In: Biodiversity and Ecosystem Function. Schulze E.D. & Mooney H.A. (eds.) Ecological Studies 99. Springer Verlag. Berlin. pp: 115-140.
Lawton, J.H. & Brown, V.K. 1993. Redundancy in Ecosystems. In: Biodiversity and Ecosystem Function. Schulze E.D. & Mooney H.A. (eds.) Ecological Studies 99. Springer Verlag. Berlin. pp: 255-270.
Levin, S.A. 1978. Pattern formation in ecological communities. In: Spatial pattern in plankton communities. Steele JA (ed) Plenum, New York. pp: 433-470.
Likens, G.E. 1992. Some applications of the ecosystems approach to environmental problems and resource management. In: Responses of Forest Ecosystems to Environmental Changes. Teller, A., Mathy, P. & Jeffers, J.N.R. (eds.). Elsevier Applied Science. pp: 16-31.
Lindeman, R.L. 1942. The trophic-dynamic aspect of ecology. Ecology 23: 399-418.
Margalef, R. 1968. Perspectives in ecological theory. University of Chicago Press, Chicago. 112p.
McNaughton, S.J. 1993. Biodiversity and Function of Grazing Ecosystems. In: Biodiversity and Ecosystem Function. Schulze E.D. & Mooney H.A. (eds.) Ecological Studies 99. Springer Verlag. Berlin. pp: 361-383.
Meyer, O. 1993. Functional groups of microorganisms. In: Biodiversity and Ecosystem Function. Schulze E.D. & Mooney H.A. (eds.) Ecological Studies 99. Springer Verlag. Berlin. pp: 67-96.
Miller, G. 1990. Living in the Environment. Wadsworth Publishing Company. Belmont, California, USA.
Moore, P.B. & Chapman, S.B. 1986. Methods in Plant Ecology. Blackwell Scientific Publications. London.
Myers, N. Mittenmeier, R.A., Mittermeier, G.G., da Fonseca, G.A.B. & Kent J. 2000. Biodiversity hotspots for conservation priorities. Nature 403: 853-858.
Naveh, 1987. Landscape ecology, management and conservation of european and levant mediterranean uplands. In: Plant Response to Stress. Tenhunen, J., Catarino, F., Lange, O., Oechel (eds.). NATO Ecological Series. Springer Verlag. Berlin.
Nicolis, G. 1991. Non-linear dynamics, self-organizing and biological complexity. In: Perspectives on biological complexity. Solbrig, O.T. Nicolis, G. (eds.) IUBS, Paris, pp: 7-49.
Nicolis, G., Prigogine, I. 1989. Exploring Complexity. An Introduction. Freenan, New York.
Odum, E. 1971. Fundamentals of Ecology, 3rd edition. Filadélfia. Saunders.
Odum, H.T. 1983. Systems ecology. An Introduction. John Wiley and Sons, N.Y.
Pimm, S.L. 1984. The Complexity and Stability of Ecosystems. Nature 307: 321-326.
Pimm, S.L. 1993. Biodiversity and the Balance of Nature. In: Biodiversity and Ecosystem Function. Schulze E.D. & Mooney H.A. (eds.) Ecological Studies 99. Springer Verlag. Berlin. pp: 347-359.
Pimm, S.L. & Raven, P.H. 2000. Extinction by numbers. Nature 403. pp: 843-845.
Raven, P.H. & Johnson, H.B. 1989. Biology. Times Mirror. Mosby College Publishing. St. Louis. Toronto.
Read, D.J. 1993. Plant-microbe Mutualisms and Community Structure. In: Biodiversity and Ecosystem Function. Schulze E.D. & Mooney H.A. (eds.) Ecological Studies 99. Springer Verlag. Berlin. pp: 181-209.
Rosén, K. 1990. Keynote paper: Measuring nutrient input to terrestrial ecosystems. In: Nutrient Cycling in Terrestrial Ecosystems. Field Methods. Application and Interpretation. Harrison, A.F., Ineson, P. & Heal, O.W. (eds.) Elsevier Applied Sciences.
Rosenzweig, C. & Parry, M.L. 1994. Potential Impact of Climate Change on World Food Supply. Nature 367: 133-138.
Sarmiento, G. 1984. Los ecosistemas y la ecosfera. Editorial Blume. Barcelona. 1ªed.
Schulze, E.D. & Mooney, H.A. 1993. Ecosystem Function of Biodiversity: A summary. In: Biodiversity and Ecosystem Function. Schulze E.D. & Mooney H.A. (eds.) Ecological Studies 99. Springer Verlag. Berlin. pp: 497-510.
Schulze, E.D. & Zwolfer, H. 1987. Introduction to the Problem of Ecosystem Analysis. In: Potentials and Limitations of Ecosystem Analysis. Schulze E.D. & Zwolfer, H. (eds.) Ecological Studies 61. Springer Verlag. Berlin. pp: 1-10.
Solbrig, O.T. 1992. Plant Traits and Adaptative Strategies. In: Biodiversity and Ecosystem Function. Schulze E.D. & Mooney H.A. (eds) Ecological Studies 99. Springer Verlag. Berlin. pp: 97-116.
Sprent, J. & Raven, J. 1985. Evolution of nitrogen-fixing symbioses. Proc. R. Soc. Edinb. B85: 215-237.
Steinberg, C.E.W. & Geller, W. 1993. Biodiversity and Interactions Within Pelagic Nutriente Cycling and Productivity. In: Biodiversity and Ecosystem Function. Schulze E.D. & Mooney, H.A. (eds.) Ecological Studies 99. Springer Verlag. Berlin. pp: 43-66.
Stevenson, F.J. & Cole, M.A. 1999. Cycles of the soil. Wiley Publishers NY.
Stubblefield, S.P., Taylor, T.N. & Trapper, J.M. 1987. Fossil mycorrhizzae: a case for symbiosis. Science 237: 59-60.
Sukachaev, V.N. 1959. The correlation between the concepts "Forest Ecosystem" and "Forest Biogeocenose" and their importante for the classification of forests. Silva Fennica 105: 94-97.
Swift, M.J. & Anderson, J.M. 1993. Biodiversity and Ecosystem Function In Agricultural Systems. In: Biodiversity and Ecosystem Function. Schulze E.D: & Mooney H.A. (eds.) Ecological Studies 99. Springer Verlag. Berlin. pp: 15-41.
Tamm, O. 1992. The evolution of ecosystem thinking in european ecology. In: Responses of Forest Ecosystems to Environmental Changes. Teller, A., Mathy, P. & Jeffers, J.N.R. (eds.) Elsevier Applied Science. pp: 5-15.
Tansley, A.G. 1935. The use and abuse of vegetational concepts and terms. Ecology 16: 284-307.
Ulrich, B. 1987. Stability, Elasticity and Resilience of Terrestrial Ecosystems with Respect to Matter Balance. In: Potentials and Limitations of Ecosystem Analysis. Schulze E.D. & Zwolfer H. (eds.) Ecological Studies 61. Springer Verlag. Berlin. pp: 11-48.
Van der Putten, wh 2000. Paltrogen-driven forest diversity. Nature 404: 232:233.

Comentários